Histology
Ganesh Athappan MD Ashley Davidoff MD
Introduction
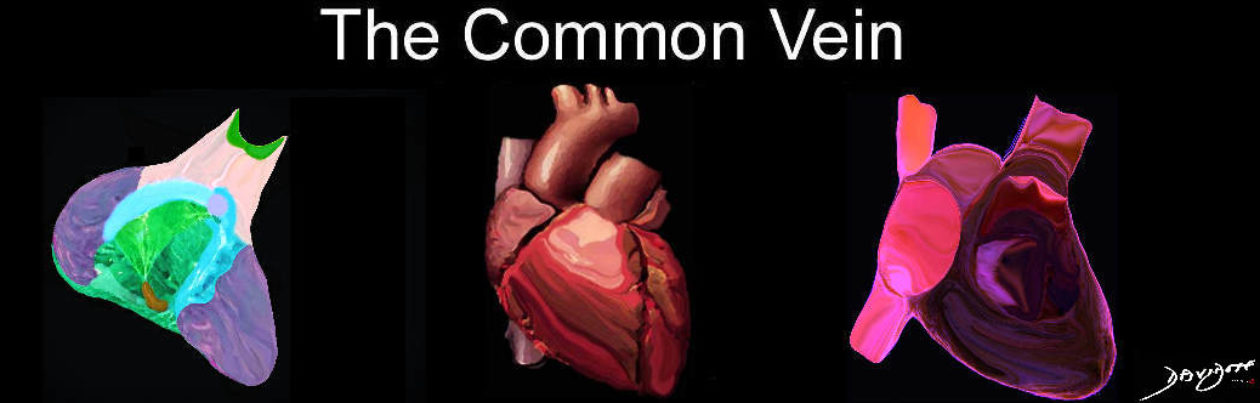
The layers of the heart conform to the basic pattern seen in the histology of other tubular structure except that the muscle layer dominates.
Tubular structures in the body have a basic structural makeup of an inner layer lined with an epithelial layer abutting the lumen, a middle functional layer and an outer protective layer or skin. In the heart the inner layer is called the endocardium, middle muscular layer is called the myocardium, and the outer layer is called the pericardium.
Ashley Davidoff, M.D. 06620c02.8s
The layers are complex in their makeup. The endocardium consists of an endothelial layer which rests on a subendothelial layer and basement membrane. Between the endocardium and myocardium there is a subendocardial layer. The myocardial layer is complex in the manner in which the myocardial cells are oriented and organized. The pericardial layer consists of two basic layers between which is the pericardial space, and each of which is made of unique tissue.

Principles
Endocardium:
The endocardium consists of a single layer of squamous endothelial cells that rests on a layer of fine collagen fibers and a basement membrane.
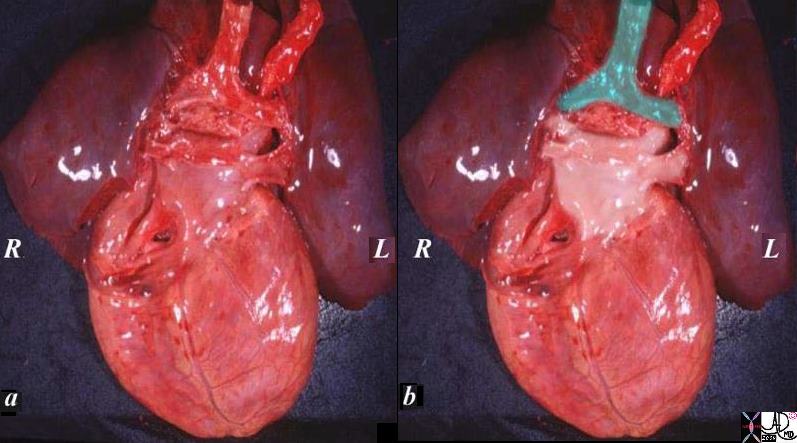
The specimen of heart and lungs is examined from posteriorly with the left atrium and 4 pulmonary veins. (overlaid in pearl white) The trachea and bronchi are intact (green). The left atrium lies just below in the angle of the carina.
Ashley Davidoff
TheCommonVein.net
08376c02.82s
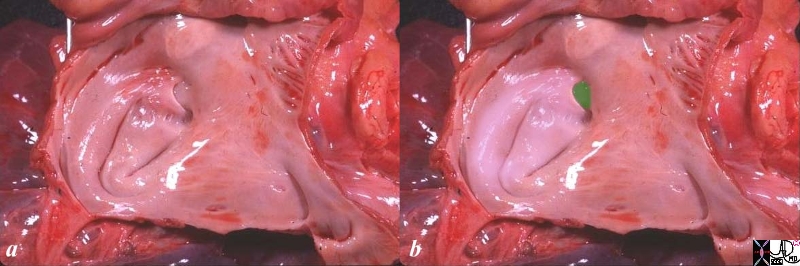
The anatomic specimen has been opened from its posterior aspect revealing a relatively smooth walled inner lining. The left atrial appendage(laa) has been opened revealing fine pectinate muscles. 5 pulmonary veins are seen entering the left atrium, with 2 entering from the left lower vein. The atrial septum with fossa ovalis and foramen ovale are smooth except for the “u” shaped upper border of the septum primum that allows identification of the left atrium. the glistening surface of the endothelium is exemplified in this image.
Ashley Davidoff
TheCommonVein.net
06426c01.81s
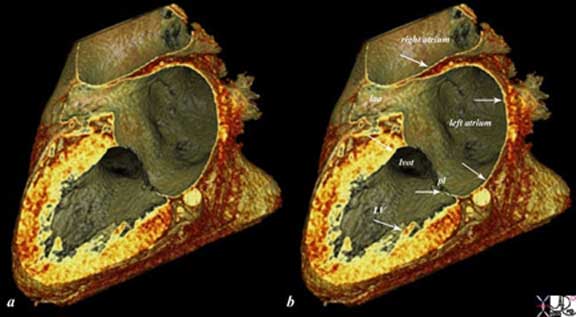
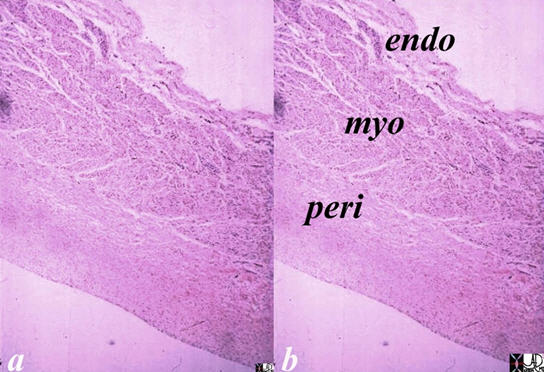
The reconstructed CT scan is in the same plane as the autopsy specimen above, and shows a distended left ventricle (LV), left atrium (LA) and right atrium (RA). The thin endocardium (white arrows) throughout the heart is one continuous sheet of tissue connected across the whole circulatory system and it is more fibrous in nature. The thin white layer is seen in the left atrium, left atrial appendage (LAA) over the posterior leaflet of the mitral valve (pl) in the left ventricle (LV) and in the right atrium. The surface of the left ventricle, left ventricular outflow tract (LVOT) and right atrium are also lined by the endothelium. Ashley Davidoff
TheCommonVein.net
47824c02.8s
The squamous endothelial cells are shaped like the scales of a fish and have oval or round nuclei. The squamous lining of the entire heart is continuous with the endothelial cells lining all the vessels in the body forming one large sheet of tissue spread throughout the entire cardiovascular system. Perhaps one can think of it as a tubular table cloth that extends from the toes to the head –one long, extensive, and remarkable sheet of tissue.
The endothelium rests on a continuous layer of fine collagen fibers, which is itself anchored by a basement membrane. These two structures (fine collagen layer and basement membrane) form the subendothelium. Beneath the subendothelium is a thicker layer of dense connective tissue forming the subendocardium. It is composed of collagen fibers , elastic fibers , smooth muscle cells , small blood vessels, and in the ventricles may contain specialized cardiac muscle cells of the conduction system . The connective tissue in this region binds the endocardium to the myocardium.
The endocardium is variable in thickness, being thickest in the atria (left > right) and thinnest in the ventricles particularly the left ventricle. Localized areas of endocardial thickening (jet lesions) are common, particularly in the atria, as a result from turbulent blood flow within the chamber.
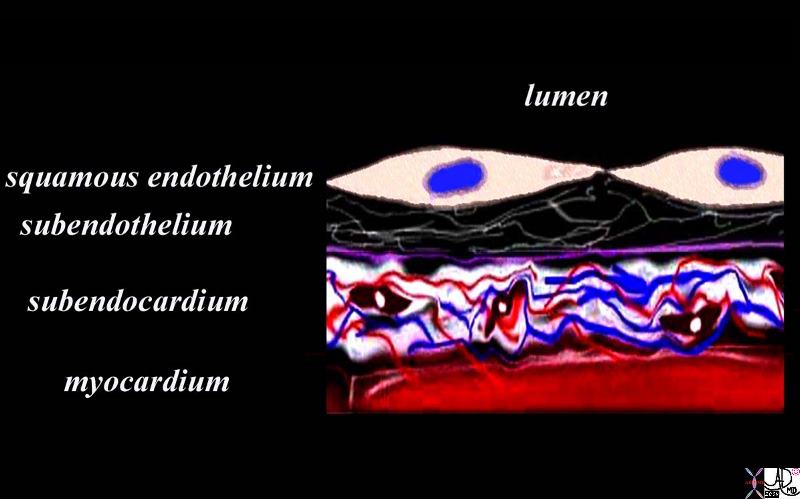
The diagram shows the endothelial layer consisting of a single layer of squamous cells abutting the cardiac lumen. Below this layer there is a fine network of collagen fibers and a basement membrane (purple) . The fine collagen layer and basement membrane together are called the subendothelial layer. The endothelium is anchored by the basement membrane. Below the basement membrane is the subendocardium which consists of dense collagen elastic tissue, capillary network, arterioles, venules, smooth muscle cells and elastic tissue. The red tissue below this layer is the myocardium.
The subendocardium is the equivalent of the submucosa that brings the lifelines of vessels and nerves to the cellular level.
Courtesy Ashley Davidoff MD Davidoff art copyright 2009 86815b14.8s
Macroscopically the endothelial surface is shiny, smooth, and moist.
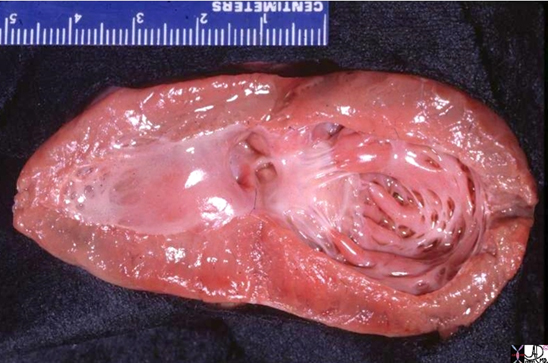
The left ventricle that has been opened along the ventricular septum shows the septal wall to your left and the free wall with the papillary muscles to your right. Note the glistening endothelium which has a whitish sheen. The myocardium lies directly under the endocardium. It is lighter red, muscular and rubbery. Note also the two sets of papillary muscles, the fibrous nature of the valves and the fibrous continuity of the mitral valve with the aortic valve. A VSD is noted on the superior aspect of the septum. Courtesy of: Ashley Davidoff, M.D.
08345b.8
The reconstructed CT scan is in the same plane as the autopsy specimen above, and shows a distended left ventricle (LV), left atrium (LA) and right atrium (RA). The thin endocardium (white arrows) throughout the heart is one continuous sheet of tissue connected across the whole circulatory system and it is more fibrous in nature. The thin white layer is seen in the left atrium, left atrial appendage (LAA) over the posterior leaflet of the mitral valve (pl) in the left ventricle (LV) and in the right atrium. The surface of the left ventricle, left ventricular outflow tract (LVOT) and right atrium are also lined by the endothelium.All images courtesy: Ashley Davidoff, M.D.
47824c02.8s
There are certain diseases that cause thickening of the endocardium usually caused by subendocardial ischemia. Endocardial fibroelastosis (EFE) which is seen in the endocardium of the left ventricle in hypoplastic left heart syndrome is shown below. The entity is thought to result from ischemic changes induced by the suprasystemic pressures in the left ventricle caused by severe left ventricular outflow obstruction and resulting in subendocardial ischemia and secondary fibrosis.
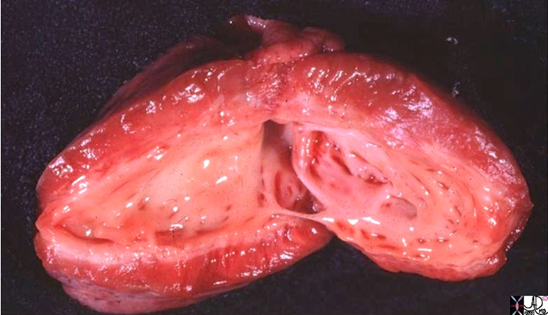
The gross pathology specimen is a case of hypoplastic left heart with aortic stenosis and mitral stenosis, revealing a thickened endocardium with left ventricular hypertrophy and deformed and thickened mitral valve and papillary muscles. The thickened endocardium is caused by the high pressures generated in the LV causing subendocardial ischemia and resulting in a condition called endocardial fibroelastosis (EFE). Images courtesy of: Ashley Davidoff, M.D. 08058.8s
Myocardium:
The myocardium is made up of layers of cardiac myocytes (75% of the total volume of the myocardium), attached to each other by connective tissue fibers, and arranged in spiral bundles. The myocardium is the contractile layer of the heart and hence the major function of myocardial muscle cells is to execute the cardiac contraction-relaxation cycle. The cell is striated, with a single nucleus, with dark staining intercalated discs. It is thickest in the left ventricle and thinnest in the atria. The overall architecture is one of muscles arranged in spiral and circular bundles.
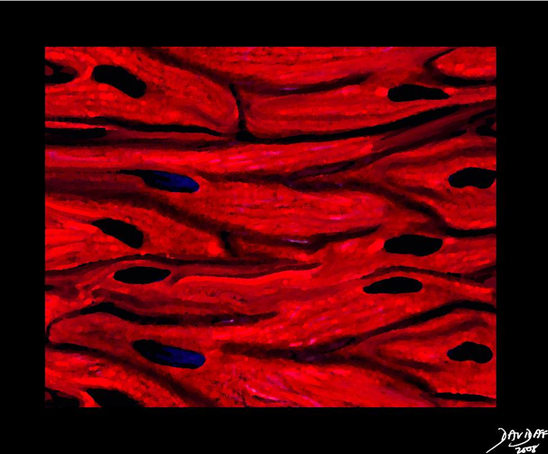
Ashley Davidoff MD
32907b03.61k.8s
|
Low Power View of the Myocardium |
|
The myocytes tend to be long, thin, and slightly rectangular. The nuclii are round and blue. Connective tissue between the muscle cells is seen as pale pink collagenous material in which parts of blood vessels can be seen. 03885b01 Courtesy Ashley Davidoff copyright 2009 |

Ventricular Myocytes
The muscles of the ventricles are arranged as
(1) external longitudinal layer
(2) middle circular and
(3) inner longitudinal layer.
Isolated bundles of cardiac muscles project into the lumen of the ventricle forming the trabeculae carnea.
The overall architecture of the myocardium of the left ventricle is in the form of a figure of 8 with one side attached to the subendocardium and then a gradual helical twist almost to 180 degrees as it extends to the subepicardial surface.
A myofiber or myocyte is the basic structural and functional unit of the myocardium. The cells are held together by surrounding collagenous connective tissue with strands of collagen also connecting the myofibers to each other. Within the myofiber there are myofibrils.
Light Microscopic Details of the Cardiac Myocyte:
The myocyte is composed of myofibrils, mitochondria, T systems, sarcoplasmic reticula, embedded within the cytoplasm.
Under the light microscope, the atrial and ventricular myocytes have cross striations and are branched and often contain two nuclii. Each myocyte is bound by a complex cell membrane, the sarcolemma and is filled with rod-like bundles of myofibrils . The latter are the contractile elements.
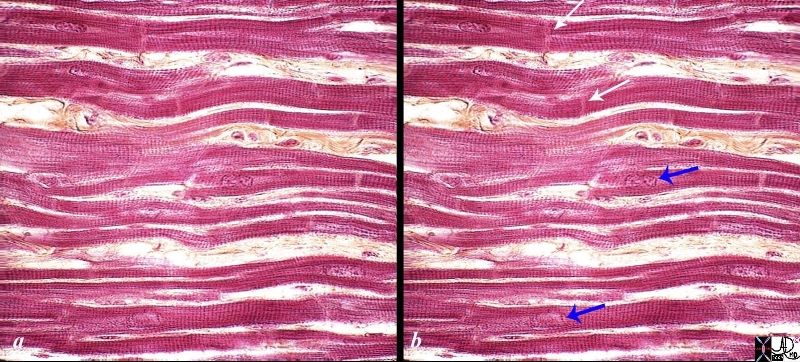
High Power of the Myocardium |
|
The histological slide shows normal myocardium with the white arrows pointing to the borders between two cells at the intercalated discs. The blue arrows point to the nuclii of the cells. The cross striations are very obvious, while the longitudinal striations are a little more difficult to visualize. Connective tissue can be seen between the cells (myocytes). 86816c02.8s Source unknown |
Characteristics of ventricular myocytes:
Size;
Length : 60-140 μm (red cell about 5 μm and liver cell about 20-30 μm)
Diameter: about 20 μm
Volume : 15,000-45,000cubic μm
Shape:
Long and narrow
Ventricular myocytes have branching bundles allowing the cells to connect both structurally and functionally as a syncitium. There is little interstitial collagen but abundant mitochondria and sarcomeres.
Character
Syncitial arrangement
Intercalated discs are prominent with end to end transmission.
T tubules are plentiful (invagination of the sarcolemma)
The sarcolemma of the myocyte invaginates the cytoplasm to form an extensive tubular network (the T tubules) that extends the extracellular space into the interior of the cell. The nucleus, which contains the cell’s genetic information, is often centrally located. Some myocytes have several nuclei. Interspersed between the myofibrils and immediately beneath the sarcolemma are many mitochondria, the main function of which is to generate the energy in the form of adenosine triphosphate (ATP) needed to maintain the heart’s contractile function and the associated ion gradients. The sarcoplasmic reticulum (SR) is a fine network spreading throughout the myocytes, demarcated by a lipid bilayer, which is similar to that of the sarcolemma. The calcium release channels (also called the ryanodine receptors) are found in the expanded parts of the SR that lie in very close and in apposition to the T tubules. These are called subsarcolemmal cisternae or the junctional SR. The second part of the SR, the longitudinal or network SR, consists of ramifying tubules and is involved with the uptake of calcium that initiates relaxation. Calcium is taken up into the SR, and is then stored at high concentration in a number of storage proteins, including calsequestrin, before being released again. The cell boundaries are seen as thin, typically dark-staining lines dividing adjacent cardiac muscle cells. They are called the intercalated discs.
The manner in which the myocytes are arranged for optimal performance is quite an ingenious trick of nature. With about 15% myocyte shortening the heart can eject between 55-65% of its load. In the mid body the cells are circumferentially arranged and are twisted one way under the subendocardium and twisted the opposite way below the epicardium. During systole this allows for shortening in both the axial plane and along the long axis as well as allowing for a wringing or twisting type action to the pattern of contraction. The base moves clockwise and the apex moves counterclockwise. This type of wringing action, (likened to the wringing action one performs to dry a wet towel) not only allows for efficient emptying but also is thought to have a vacuum effect as relaxation occurs.
Atrial Myocytes:
The muscles of the atria extend as an arc from the front to back and partly diagonally across the atria from one side to the other . Bundles of atrial tissue are separated by wide areas of collagen. Prominent bundles of atrial muscles form the pectinate muscles. Atrial myocytes are the basic structural and functional unit of the atrium. The characteristics of atrial myocytes are slightly different to the ventricular myocytes:
Size
In general the atrial myocyte is much shorter, thinner, and less voluminous than the ventricular myocyte. T tubules are rare whereas in ventricular muscle they are abundant.
Length : 20 μm
Diameter: 5-6 μm
Volume : approx 500 cubic μm
Shape:
Elliptical
Character
T tubules are rare
Intercalated discs with side to side and end to end transmission.
Some myocytes in the RA and fewer in the LA have neurosecretory granules for atrial natriuretic peptide.
Pericardium:
The pericardium consists of two basic layers. There is an inner layer called the visceral pericardium that is made of a layer of fibroelastic connective tissue blood vessels, lymphatics, nerve fibres, and a variable amount of adipose tissue that merges with the myocardium. The second layer called the parietal pericardium is more superficial layer, and is made up of a tougher fibroelastic layer called the fibrous pericardium and a more delicate mesothelial serous lining.
Visceral Pericardium
The epicardium (or visceral pericardium) forms the outer covering of the heart. (Cormack). Its free surface is composed of a single layer of flat to cuboidal mesothelial cells resting on a layer of connective tissue rich in elastic fibers. The subepicardial space adjacent to the myocardium contains blood vessels, nerves and abundance of fat cells.
Parietal pericardium
The parietal pericardium has two parts; an inner portion that has a serous lining and an outer fibrous component. The latter is made from collagen with a small amount of elastin. The collagen is organized in criss-cross pattern.
When the inner single cell thick cuboidal mesothelial surface of the linings of both the parietal and visceral pericardium are reviewed under the electron microscope they reveal microvilli and long cilia which secrete fluid and provide a low friction surface.
Below the mesothelial epithelia a layer of elastic fibrils supports the epithelium.
There is a rich network of lymphatics that provide the substrate for the pericardial fluid.
Fibrous Skeleton:
The fibrous skeleton of the heart , formed of connective tissue, separates the atria from the ventricles and includes the annuli fibrosi – (rings of dense connective tissue around the valves), trigona fibrosi ( masses of fibrous connective tissue connecting the annuli) and the septum membranicum (membranous spot at the top of the interventricular septum). It is composed of dense collagen fibers, scattered elastic fibers and occasional fat cells. (less regular than tendon, more regular than dermis). The cardiac skeleton serves as the place of origin and attachment of muscles of the atria and ventricles.
Valve Leaflets
Valve leaflets are composed of three layers (ventricularis, fibrosa and spongiosa) and two general cell types (1) endothelium – non thrombogenic external lining and valvular interstitial cells – heterogeneous cell population composed of fibroblast and smooth muscle cells. The cardiac valves are lined with endothelium and all have a similar three- layered architecture consisting predominantly of a dense collagenous core, the fibrosa layer, close to the outflow surface and continuous with the valvular supporting structures. Towards the ventricular/atrial cavity the ventricualis/atrialis layer is found and it is rich in elastin. Between these two layers is the centrally located core of loose connective tissue, the spongiosa layer. The collagen of the fibrosa layer is responsible for the mechanical strength of the valve, whereas the connective tissue of the spongiosa layer works as a shock-absorber. Normal leaflets and cusps have only scant blood vessels and are avascular structures.
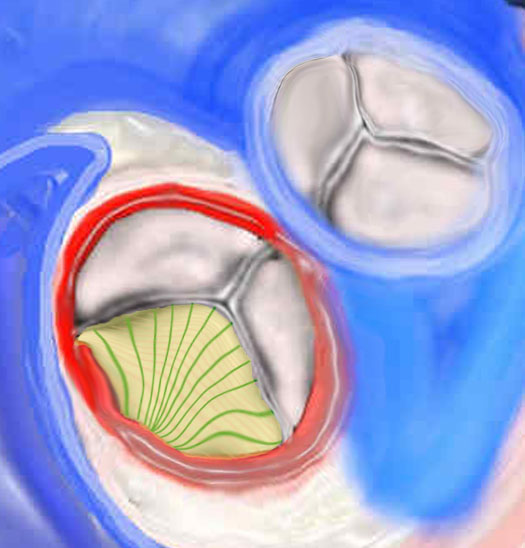
|
Histological Makeup of the Aortic Valve |
|
There are 3 layers of collagen within the valve. The first image (top) shows the fibrosa layer on the aortic side of the valve. (pink) This layer consists of radially oriented fibers extending from commissure to commissure. It has corrugated form enabling it to stretch during systole. The second middle layer is called the spongiosa green and its fibers extend at right angles to the fibers of the fibrosa. On the ventricular side in the third image a cross section of the histological makeup of the aortic valve is shown with the upper layer being on the aortic side and the yellow layer of ventricularis is shown. The ventricularis is a sheet of tissue called. (yellow) 47682d07 47682d03 47682eb06.800 Copyright 2009 Courtesy Ashley Davidoff Davidoff art |

Histology of the Cardiac Conduction System :
The conduction system is made of myocardial fibers designed for rhythm rather than contraction. They include the nodal cells ( AV nodal , SA nodal) and the AV bundle along with its branches (Purkinje cells).
Nodal Cells: Nodal cells are smaller than ordinary cardiac myocytes, contain fewer and poorly organized myofibrils and no intercalated discs.
Purkinje cells: They are specialized myocytes found in the subendocardium of the ventricles. Purkinje cells are rich in glycogen and mitochondria and often have two more nuclei centrally placed.
Charactersitics of Purkinje cells:
Size :
length 150-200μm
diameter : 35– 40μm
Volume : 135,000-250,000 cubic μm
Shape :
Long and broad , lighter staining and with abundant sarcoplasm and scarce sarcomere
Character
T- Tubules are absent
Intercalated discs are very prominent with abundant gap junctions for fast end-to-end transmission.